
Analysis of Grass
Background on Grass



Celignis founder Daniel Hayes has considerable experience in the chemical and near-infrared analysis of grasses and has characterised a large number of samples from a wide variety of species.
Components of Grasses
The above ground shoot component of grasses consists of a stem and leaves. The stems are mostly hollow, cylindrical and interrupted at intervals by swollen joints or nodes from which the leaves originate. The parts of the stem between the nodes are termed the internodes.
Rhizomes are a special type of shoot that occur in some grassy species (e.g. miscanthus). They grow underground in a horizontal manner and their internodes are usually short. The only leaves present on rhizomes are small and scale-like. Rhizomes serve as survival organs after periods of senescence and also are often used for storage.
The lower portion of the leaf forms a sheath, which protects the shoots. The second half of the leaf opens out to the leaf blade.
The proportion of biomass components vary with the species. For example, with barley and wheat straw, internodes dominate (59% and 73% of the dry mass, respectively) while leaves (including sheath and blade) constitute an intermediate, and nodes a minor portion of the aboveground dry matter.
Extractives tend to be more prevalent in grasses than woody biomass. In particular, concentrations of water-soluble carbohydrates tend to be relatively high. Glucose and fructose are the two most important free-monosaccharides that occur in grasses and can have dry matter concentrations of approximately 1-3%. More prevalent is the disaccharide sucrose, which can take up 2-8% of the dry matter in forage grasses. In certain grasses, such as perennial ryegrass (Lolium perenne) and cocksfoot (Dactylis glomerata), oligosaccharides such as raffinose and stachyose have been detected, but only in relatively small concentrations. The polysaccharide fructans tend to be the most abundant of the water soluble carbohydrates, constituting around 5-9% of the dry matter of forage grasses, although values as high as 12% have been reported for perennial ryegrass.
Analysis of Grass at Celignis
Celignis Analytical can determine the following properties of Grass samples:
Lignocellulosic Properties of Grass
Cellulose Content of Grass
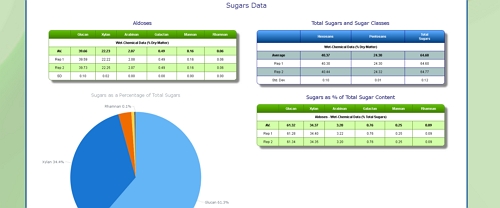
Click here to see the Celignis Analysis Packages that determine Cellulose Content
Request a QuoteCellulose Content
Hemicellulose Content of Grass
Click here to see the Celignis Analysis Packages that determine Hemicellulose Content
Request a QuoteHemicellulose Content

Lignin Content of Grass

Click here to see the Celignis Analysis Packages that determine Lignin Content
Request a QuoteLignin Content
Starch Content of Grass
Click here to see the Celignis Analysis Packages that determine Starch Content
Request a QuoteStarch Content

Uronic Acid Content of Grass

Click here to see the Celignis Analysis Packages that determine Uronic Acid Content
Request a QuoteUronic Acid Content
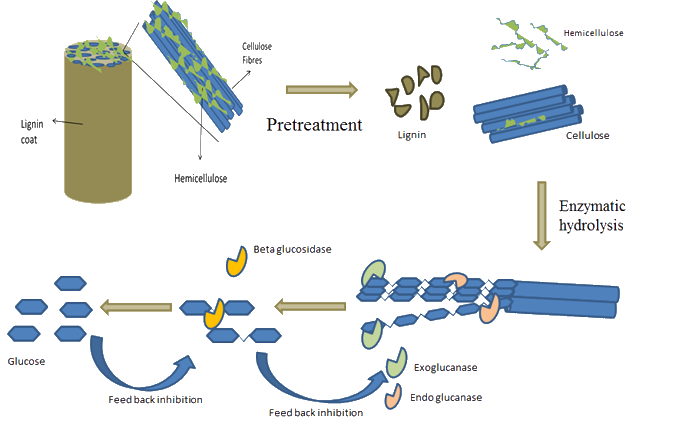
Enzymatic Hydrolysis of Grass
Click here to see the Celignis Analysis Packages that determine Enzymatic Hydrolysis
Request a QuoteEnzymatic Hydrolysis
Bioenergy Properties of Grass
Ash Content of Grass

Click here to see the Celignis Analysis Packages that determine Ash Content
Request a QuoteAsh Content
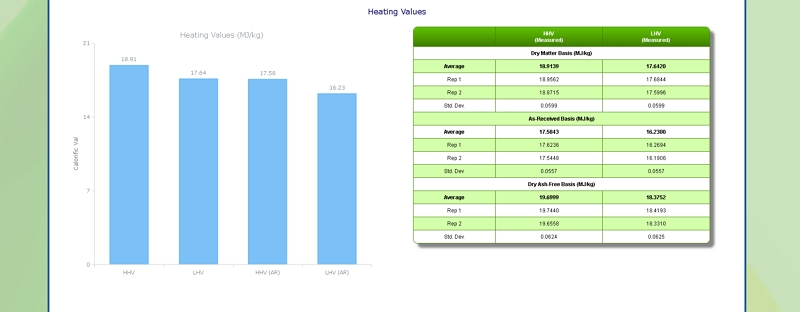
Heating (Calorific) Value of Grass
Click here to see the Celignis Analysis Packages that determine Heating (Calorific) Value
Request a QuoteHeating (Calorific) Value
Ash Melting Behaviour of Grass
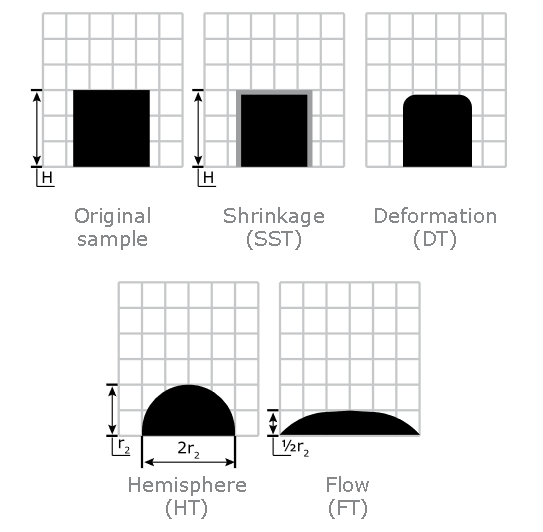
Ash Shrinkage Starting Temperature (SST) - This occurs when the area of the test piece of Grass ash falls below 95% of the original test piece area.
Ash Deformation Temperature (DT) - The temperature at which the first signs of rounding of the edges of the test piece occurs due to melting.
Ash Hemisphere Temperature (HT) - When the test piece of Grass ash forms a hemisphere (i.e. the height becomes equal to half the base diameter).
Ash Flow Temperature (FT) - The temperature at which the Grass ash is spread out over the supporting tile in a layer, the height of which is half of the test piece at the hemisphere temperature.
Click here to see the Celignis Analysis Packages that determine Ash Melting Behaviour
Request a QuoteAsh Melting Behaviour
Major and Minor Elements in Grass
We can also determine the levels of 13 different minor elements (such as arsenic, copper, and zinc) that may be present in Grass.
Click here to see the Celignis Analysis Packages that determine Major and Minor Elements
Request a QuoteMajor and Minor Elements

Analysis of Grass for Anaerobic Digestion
Biomethane potential (BMP) of Grass

At Celignis we can provide you with crucial data on feedstock suitability for AD as well as on the composition of process residues. For example, we can determine the biomethane potential (BMP) of Grass. The BMP can be considered to be the experimental theoretical maximum amount of methane produced from a feedstock. We moniotor the volume of biogas produced allowing for a cumulative plot over time, accessed via the Celignis Database. Our BMP packages also involve routine analysis of biogas composition (biomethane, carbon dioxide, hydrogen sulphide, ammonia, oxygen). We also provide detailed analysis of the digestate, the residue that remains after a sample has been digested. Our expertise in lignocellulosic analysis can allow for detailed insight regarding the fate of the different biogenic polymers during digestion.
Click here to see the Celignis Analysis Packages that determine BMP
Request a QuoteBMP
Physical Properties of Grass
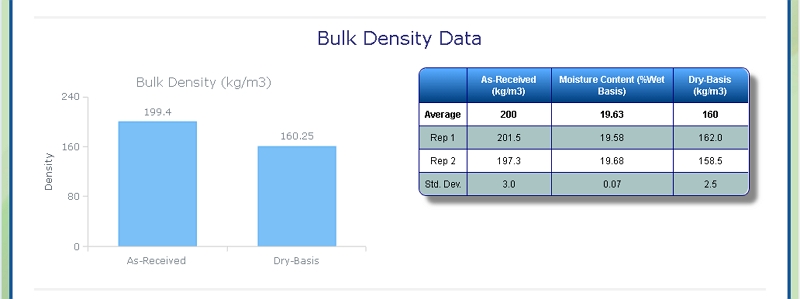
Bulk Density of Grass
At Celignis we can determine the bulk density of biomass samples, including Grass, according to ISO standard 17828 (2015). This method requires the biomass to be in an appropriate form (chips or powder) for density determination.
Click here to see the Celignis Analysis Packages that determine Bulk Density
Request a QuoteBulk Density
Particle Size of Grass
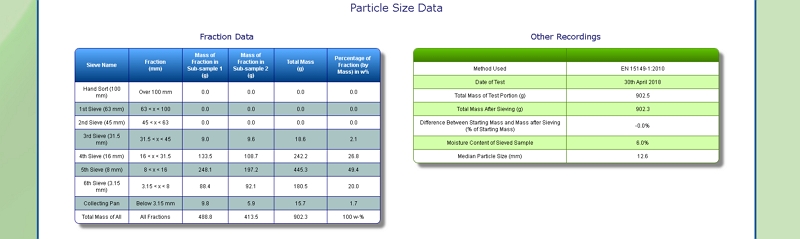
Our lab is equipped with a Retsch AS 400 sieve shaker. It can accommodate sieves of up to 40 cm diameter, corresponding to a surface area of 1256 square centimetres. This allows us to determine the particle size distribution of a range of samples, including Grass, by following European Standard methods EN 15149- 1:2010 and EN 15149-2:2010.
Click here to see the Celignis Analysis Packages that determine Particle Size
Request a QuoteParticle Size
Next-generation biofuels from renewable sources have gained interest among research investigators, industrialists, and governments due to major concerns on the volatility of oil prices, climate change, and depletion of oil reserves. Biobutanol has drawn signicant attention as an alternative transportation fuel due to its superior fuel properties over ethanol. e advantages of butanol are its high energy content, better blending with gasoline, less hydroscopic nature, lower volatility, direct use in convention engines, low corrosiveness, etc. Butanol production through (acetone, butanol, and ethanol) ABE fermentation is a well-established process, but it has several drawbacks like feedstock cost, strain degeneration, product toxicity, and low product concentrations. Lignocellulosic biomass is considered as the most abundant, renewable, low-cost feedstock for biofuels. Production of butanol from lignocellulosic biomass is more complicated due to the recalcitrance of feedstock and inhibitors generated during the pretreatment and hydrolysis process. Advanced fermentation and product recovery techniques are being researched to make biobutanol industrially viable. | |
Biomass feedstock having less competition with food crops are desirable for bio-ethanol production and such resources may not be localized geographically. A distributed production strategy is therefore more suitable for feedstock like water hyacinth with a decentralized availability. In this study, we have demonstrated the suitability of this feedstock for production of fermentable sugars using cellulases produced on site. Testing of acid and alkali pretreatment methods indicated that alkali pretreatment was more efficient in making the sample susceptible to enzyme hydrolysis. Cellulase and ?-glucosidase loading and the effect of surfactants were studied and optimized to improve saccharification. Redesigning of enzyme blends resulted in an improvement of saccharification from 57% to 71%. A crude trial on fermentation of the enzymatic hydrolysate using the common baker�s yeast Saccharomyces cerevisiae yielded an ethanol concentration of 4.4 g/L. | |
Examples of Other Feedstocks Analysed at Celignis
Lignin (Klason), Lignin (Acid Soluble), Acid Insoluble Residue, Ash (Acid Insoluble),
As P10 plus protein-corrected lignin, water-soluble sugars, uronic acids, acetyl content and starch.
Glucuronic Acid, Galacturonic Acid, Mannuronic Acid, Guluronic Acid, 4-O-Methyl-D-Glucuronic Acid, Iduronic Acid,
Total Sugars in Enzyme Hydrolysate, Glucose in Enzyme Hydrolysate, Xylose in Enzyme Hydrolysate, Arabinose in Enzyme Hydrolysate, Mannose in Enzyme Hydrolysate, Galactose in Enzyme Hydrolysate, Rhamnose in Enzyme Hydrolysate, Cellobiose in Enzyme Hydrolysate, Enzymatic Hydrolysis Kinetics, Cellulose Conversion Yield, Xylan Conversion Yield, Combined Sugar Yield, Cellulose Conversion Rate, Xylan Conversion Rate,
As P121 plus comparisons with data from the non-pretreated original sample, including: Increase in Cellulose Accessibility after Pre-Treatment, Percent Increase in Cellulose Conversion Efficiency, Percent Increase in Cellulose Conversion Rate.
As P9 but on the solid residue after enzymatic hydrolysis.
Formic Acid, Acetic Acid, Levulinic Acid, Furfural, Hydroxymethylfurfural,
Includes all hydrolysate sugars and kinetics in P121 and: Cellulose Conversion Yield, Cellulose Conversion Rate
Includes all hydrolysate sugars and kinetics in P121 and: Xylan Conversion Yield, Xylan Conversion Rate
Total Sugars in Enzyme Hydrolysate, Glucose in Enzyme Hydrolysate, Maltose in Enzyme Hydrolysate, a-Amylase Hydrolysis Kinetics, Glucoamylase Hydrolysis Kinetics,
Glucose, Xylose, Fructose, Sucrose, Mannose, Arabinose, Galactose, Rhamnose, Xylitol, Sorbitol, Trehalose, Mannitol, Arabinitol, Glycerol, Raffinose,
Levulinic Acid, Formic Acid, Hydroxymethylfurfural, Furfural, Acetic Acid, gamma-Valerolactone,
Xylobiose, Xylotriose, Arabinobiose, Arabinotriose,
Maltose, Maltotriose, Maltotetraose, Maltopentaose, Maltohexaose, Maltoheptaose, Maltooctaose,
Glucuronic Acid, Galacturonic Acid, Mannuronic Acid, Guluronic Acid, 4-O-Methyl-D-Glucuronic Acid, Iduronic Acid,
Gibberellic Acid, Indole-3-acetic acid, Indole-2-acetic acid, Indole-3-propionic acid, Indole-3-butyric acid, 6-Benzylaminopurine, Kinetin riboside, Abscisic acid, Salicylic acid,
beta-Carotene, Ergocalciferol (Vitamin D2), Alpha-tocopherol (vitamin E), Phylloquinone (Vitamin K1),
Thiamine (Vitamin B1), Riboflavin (Vitamin B2), Niacin (Vitamin B3), Niacinamide (vitamin B3), Pantothenic Acid (Vitamin B5), Pyridoxine (Vitamin B6), Folate (Vitamin B9), Cobalamin (Vitamin B12), Ascorbic Acid (Vitamin C),
Fucose, Mannitol, Glucose, Xylose, Mannose, Arabinose, Galactose, Rhamnose, Total Sugars, Glucuronic Acid, Galacturonic Acid, Mannuronic Acid, Guluronic Acid, Iduronic Acid,
Alanine, Arginine, Aspartic Acid, Asparagine, Cystine, Glutamic Acid, Glutamine, Glycine, Histidine, Isoleucine, Leucine, Lysine, Methionine, Phenylalanine, Proline, Serine, Threonine, Tryptophan, Tyrosine, Valine,
Aluminium, Calcium, Iron, Magnesium, Phosphorus, Potassium, Silicon, Sodium, Titanium,
Arachidic Acid, Behenic Acid, Decanoic Acid, Erucic Acid, Lauric Acid, Linoleic Acid, Linolenic Acid, Myristic Acid, Caprylic Acid, Oleic Acid, Palmitic Acid, Palmitoleic Acid, Stearic Acid, Lignoceric Acid,
Fucoxanthin, Astaxanthin, Chlorophyll-c, Chlorophyll-a, Chlorophyll-b, Lutein, beta-Carotene, Neoxanthin, Antheraxanthin, Violaxanthin,
Gibberellic Acid, Indole-3-acetic acid, Indole-2-acetic acid, Indole-3-propionic acid, Indole-3-butyric acid, 6-Benzylaminopurine, Kinetin riboside, Abscisic acid, Salicylic acid,
beta-Carotene, Ergocalciferol (Vitamin D2), Alpha-tocopherol (vitamin E), Phylloquinone (Vitamin K1),
Thiamine (Vitamin B1), Riboflavin (Vitamin B2), Niacin (Vitamin B3), Niacinamide (vitamin B3), Pantothenic Acid (Vitamin B5), Pyridoxine (Vitamin B6), Folate (Vitamin B9), Cobalamin (Vitamin B12), Ascorbic Acid (Vitamin C),
Chlorophyll-a, Chlorophyll-b, Lutein, beta-Carotene, Neoxanthin, Astaxanthin, Zeaxanthin, Antheraxanthin, Violaxanthin,
Biomethane Potential (BMP), Total Biogas Volume, Total Solids, Volatile Solids, pH, Biogas Methane Content, Biogas Carbon Dioxide Content, Biogas Oxygen Content, Biogas Hydrogen Sulphide Content, Biogas Ammonia Content,
Total Solids, Volatile Solids, pH, Chemical Oxygen Demand (COD), Phosphorus, Potassium, Ammonia, Carbon, Hydrogen, Nitrogen, Sulphur,
Total Solids, Volatile Solids, pH, Chemical Oxygen Demand (COD), Phosphorus, Potassium, Ammonia, Carbon, Hydrogen, Nitrogen, Sulphur,
Residual Biogas Potential (RBP), Total Biogas Volume, Total Solids, Volatile Solids, pH, Biogas Methane Content, Biogas Carbon Dioxide Content, Biogas Oxygen Content, Biogas Hydrogen Sulphide Content, Biogas Ammonia Content,
Acetic Acid, Lactic Acid, Propionic Acid, Butyric Acid, Isobutyric Acid, Valeric Acid, Isovaleric Acid,
Levoglucosan, Cellobiosan, Mannosan, Galactosan, Glucose, Xylose, Mannose, Arabinose, Galactose, Rhamnose, Fucose, Sucrose, Cellobiose, Total Sugars,
31 constituents including Phenol, Furfural, Syringol, and Vanillin
Specific Surface Area (Nitrogen Gas Adsorption), BET Isotherm (5 Point Using Nitrogen),
Specific Surface Area (Nitrogen Gas Adsorption), BET Isotherm (20 Point Using Nitrogen), Pore Volume (Using Nitrogen), Pore Size Distribution (Using Nitrogen), Average Pore Width (Using Nitrogen),
Specific Surface Area (Nitrogen Gas Adsorption), BET Isotherm (40 Point Using Nitrogen), Pore Volume (Using Nitrogen), Pore Size Distribution (Using Nitrogen), Average Pore Width (Using Nitrogen),
Gross Calorific Value, Net Calorific Value, Ash, Carbon, Hydrogen, Nitrogen, Sulphur, Oxygen,
Aluminium, Calcium, Iron, Magnesium, Phosphorus, Potassium, Silicon, Sodium, Titanium,
Antimony, Arsenic, Cadmium, Chromium, Cobalt, Copper, Lead, Manganese, Mercury, Molybdenum, Nickel, Vanadium, Zinc,
Ash Shrinkage Starting Temperature (Reducing), Ash Deformation Temperature (Reducing), Ash Hemisphere Temperature (Reducing), Ash Flow Temperature (Reducing),
Moisture, Ash Content (815C), Carbon, Hydrogen, Nitrogen, Sulphur, Oxygen, Chlorine, Volatile Matter, Fixed Carbon, Aluminium, Calcium, Iron, Magnesium, Phosphorus, Potassium, Silicon, Sodium, Titanium, Gross Calorific Value, Net Calorific Value, Ash Shrinkage Starting Temperature (Reducing), Ash Deformation Temperature (Reducing), Ash Hemisphere Temperature (Reducing), Ash Flow Temperature (Reducing),
As P393 plus inorganic carbon, organic carbon, TGA (under nitrogen and air), and inherent moisture
Aluminium, Calcium, Iron, Magnesium, Phosphorus, Potassium, Silicon, Sodium, Titanium,
Antimony, Arsenic, Cadmium, Chromium, Cobalt, Copper, Lead, Manganese, Mercury, Molybdenum, Nickel, Vanadium, Zinc,
Acenaphthene, Acenaphthylene, Anthracene, Benz[a]anthracene, Benzo[b]fluoranthene, Benzo[k]fluoranthene, Benzo[ghi]perylene, Benzo[a]pyrene, Chrysene, Dibenz[a,h]anthracene, Fluoranthene, Fluorene, Indeno[1,2,3-cd]pyrene, 1-Methylnaphthalene, 2-Methylnaphthalene, Naphthalene, Phenanthrene, Pyrene,
Time to Germination, Mean Shoot Length (Week 1), Mean Shoot Length (Week 2), Mean Shoot Length (Week 3), Mean Shoot Length (Week 4), Shoot Weight (Week 4), Mean Root Length (Week 4), Root Weight (Week 4),
As Deluxe package plus P383, SEM Imaging (P387) and Plant Growth Trials (P388)
Includes everything from P391 (Physical Properties Ultimate), P394 (Thermal Properties Ultimate), and P397 (Soil Amendment Ultimate)
Gross Calorific Value, Net Calorific Value, Ash, Carbon, Hydrogen, Nitrogen, Sulphur, Oxygen,
Aluminium, Calcium, Iron, Magnesium, Phosphorus, Potassium, Silicon, Sodium, Titanium,
Antimony, Arsenic, Cadmium, Chromium, Cobalt, Copper, Lead, Manganese, Mercury, Molybdenum, Nickel, Vanadium, Zinc,
Volatile Matter, Fixed Carbon, Moisture, Ash, Carbon, Hydrogen, Nitrogen, Sulphur, Oxygen, Gross Calorific Value, Net Calorific Value, Chlorine,
Ash Shrinkage Starting Temperature (Oxidising), Ash Deformation Temperature (Oxidising), Ash Hemisphere Temperature (Oxidising), Ash Flow Temperature (Oxidising),
Our Custom Chatbot is Integrated into the Celignis Database
We are proud to announce the launch of Celignis InSightAI, a powerful artificial intelligence (AI) tool developed by Celignis and integrated with the Celignis Database. It functions as a highly-customised chatbot, focused on providing insights and conclusions from the data displayed on the page. Once launched, and authorised by the user, it has access to the results presented on the Database page and the user can ask it questions about those results.
A 30-second video outlining some of its functionality is provided below.
Read...

Iowa-based Facility will Offer Our Services to US-Based Clients
We're delighted to announce that today marks the official launch date of Celignis USA. This latest development in the extension of the Celignis family will offer all of Celignis's services to US-based clients. The 10,000 sq. ft facility is located in Waterloo, Iowa and leverages Celignis's 11 years of expertise in the bioeconomy.

Click here to read more about Celignis USA and our history leading to this point.

Read...
Articles cover bioprocess development as well as analytical services across many different sectors
We currently offer over 150 analysis packages, covering close to 400 different analytes. We have been proud to provide data to a diverse array of clients over the years, including many from research institutions. A recent literature search found a large number of peer-reviewed articles citing data provided by Celignis. These papers cover our
Read...
